Тема 2: ВОЗРАСТНЫЕ ОСОБЕННОСТИ МЫШЦ
У новорожденных мышцы развиты относительно хорошо и составляют 20-22% от общей массы тела, у детей 1-2 лет 16,6%. В 6 лет масса скелетных мышц достигает 21,7%, затем она увеличивается до 33% от массы тела у женщин и 36% у мужчин. Мышечные волокна в пучках лежат рыхло, толщина их небольшая — в большинстве мышц от 4 до 22 мкм. Сухожилия развиты слабо. В дальнейшем рост мышц происходит неравномерно в зависимости от их функциональной активности, как за счет утолщения имеющихся волокон, так и путем образования новых. В первые годы жизни быстро растут мышцы верхней и нижней конечностей и их сухожилия. В период от 2 до 4 лет отмечается усиленный рост длины мышц спины и большой ягодичной мышцы. Мышцы, обеспечивающие вертикальное положение тела (в статике и передвижении), интенсивно увеличиваются с 7 лет и, особенно у подростков, с 12 до 16 лет. Поперечные размеры мышечных волокон к 18 годам достигают 20-30 мкм.
Фасции у новорожденных тонкие, рыхлые, от мышц отделяются легко. Формирование фасций начинается с первых месяцев жизни ребенка и взаимосвязано с функциональной активностью мышц.
Мы ш ц ы г о л о в ы. У новорожденных мимические мышцы развиты слабо, за исключением круговой мышцы рта и щечной; обеспечивающих акт сосания. Лобное и затылочное брюшко надчерепной мышцы выражены сравнительно хорошо, хотя сухожильный шлем развит недостаточно и рыхло соединен с надкостницей костей крыши черепа, что способствует образованию гематом при родовых травмах. Так же недостаточно сформированы жевательные мышцы, интенсивное их развитие отмечается в первые годы жизни (период прорезывания молочных зубов, особенно коренных). В этот период появляются сравнительно большие скопления жировой клетчатки между поверхностным и глубоким листками височной фасции над скуловой дугой, между височной фасцией и височной мышцей, а также между последней и надкостницей, что придает голове новорожденных и детей первых лет жизни округлые очертания. К 5-8 годам мышцы головы и их фасции хорошо развиты.
Мы ш ц ы ш е и у новорожденных и их сухожильные части развиты слабо. Лучше контурируется грудино-ключично-сосцевидная, двубрюшная и лестничные мышцы. К 5-7 годам все мышцы хорошо развиты, в 10-14 лет мускулатура шеи мало отличается от взрослого человека. Окончательного развития мышцы достигают к 20-25 годам.
Шея у ребенка относительно короткая вследствие высокого стояния грудной клетки, поэтому у новорожденных и детей до 2-3 лет треугольники шеи находятся выше, чем у взрослых.
В связи с этим меняется ориентация сосудисто-нервных образований. Характерное для взрослых людей положение треугольники шеи занимают после 15 лет.
Пластинки шейных фасций новорожденных тонкие, рыхлые, поэтому межфасциальных пространства легко сообщаются. В межфасциальных пространствах мало клетчатки, количество ее заметно возрастает лишь к 6-7 годам и достигает окончательного развития к периоду половой зрелости.
Мы ш ц ы с п и н ы развиты слабо, особенно глубокие. Они имеют значительно большую контрактильную часть, чем сухожильную. Волокна широчайшей мышцы тесно прилежат к наружной косой мышце живота, так что поясничный треугольник едва намечен. Усиленный рост ‘всех мышц спины отмечается с 2 до 4 лет, в 5-6 лет и в период полового созревания.
Мы ш ц ы г р у д и. Мускулатура грудной клетки у детей первых лет жизни слабо развита, особенно глубокие мышцы. Они хорошо контурируются в 5-6 лет, усиленно paстут в 10-12 лет (период второго детства). Наибольшие возрастные особенности присущи диафрагме, она у новорожденного хорошо развита, вес составляет 5,3% всей мускулатуры (у взрослых 1,02-1,34 %). Это связано с первостепенным ее значением в акте дыхания, так как межреберные мышцы слабо развиты. У новорожденных и детей до 5 лет диафрагма расположена высоко, что связано с горизонтальным положением ребер. Купол диафрагмы выпуклый, хорошо развит ее поясничный отдел. Грудинно-реберные и пояснично-реберные треугольники относительно больше, чем у взрослых. Сухожильная часть занимает 12-15% ее площади. К 7 годам диафрагма приобретает положение как у взрослого человека.
Мы ш ц ы ж и в о т а у новорожденных пропорционально длиннее, чем у взрослых, поскольку волокна этих мышц удлиняются за счет давления внутренних органов. У новорожденных они развиты недостаточно, поэтому не выражен рельеф передней брюшной стенки; апоневрозы мышц нежные, широкие. Мышечные слои с трудом отделяются друг от друга, так как фасции, покрывающие мышцы, слабо развиты. Переход мышечной части в апоневроз не выражен. Мышечная часть наружной косой мышцы живота относительно короче, а нижние пучки внутренней косой мышцы развиты лучше, чем у взрослых. Сухожильные перемычки прямой мышцы живота расположены высоко и в раннем детском возрасте не всегда симметричны на обеих сторонах. Влагалище прямой мышцы живота имеет обычный принцип строения. В нем слабо развита задняя стенка. Белая линия живота четко выражена, ширина ее у мечевидного отростка 558 мм, на уровне пупка 12-16 мм, особенно широка в ‘местах слияния с сухожильными перемычками прямых мышц. В верхней части белой линии и в области пупка в ней обнаруживаются истонченные участки.
Паховый канал короткий, широкий (10-15 мм). Поверхностное паховое кольцо (диаметр 0,7-1,4 см) ограничено медиальной и латеральной ножками апоневроза наружной косой мышцы живота. Медиальная ножка развита слабее латеральной, межножковые волокна отсутствуют, они видны лишь со 2-го года жизни ребенка. Поперечная фасция тонкая, предбрюшинного скопления клетчатки почти нет. Глубокое паховое кольцо канала в виде воронкообразного углубления в поперечной фасции, прикрытое брюшиной, его диаметр. 2-4 мм. Паховый канал окончательно формируется к 3 годам. Пупочное кольцо новорожденных расположено относительно низко, нижняя его часть укреплена соединительной тканью, верхний отдел слабее нижнего и нередко является местом возникновения пупочной грыжи.
Интенсивное нарастание мышечной массы, укрепление апоневрозов, уплотнение фасций наблюдается в период начала ходьбы ребенка (1-3 года).
Мы ш ц ы к о н е ч н о с т е й развиты слабо. Из особенностей мышц конечностей новорожденных следует отметить значительную длину сократительной части, благодаря чему объем конечностей (особенно предплечья и голени) в проксимальном и дистальном отделах почти одинаков. У взрослых же в нижней трети предплечья и голени практически расположены только сухожилия мышц. Мышцы глубоких слоев дифференцированы нечетко, часто представлены общим мышечным пластом. Мышцы верхней конечности составляют 27% всей мышечной массы, а мышцы нижней конечности — 38 %, в то время как у взрослого соответственно — 28 % и 54 % .
у новорожденных имеется ряд особенностей топографии верхней и нижней конечностей. Бедренный канал — его внутреннее отверстие широкое, длина канала мала. Наружное отверстие также широкое (овальная ямка), расположено сразу под паховой связкой, заполнено рыхлой клетчаткой. Мышечная и сосудистая лакуны у новорожденных относительно шире и расположены более вертикально, чем у взрослых, в связи с воронкообразной формой таза.
Костно-фиброзные каналы и синовиальные влагалища КИСТИ’ и стопы сформированы. Из особенностей их строения следует отметить, что у новорожденных синовиальные влагалища мизинца и большого пальца верхней конечности не сообщаются с общим синовиальным влагалищем запястья, связь формируется в течение 1-го года жизни.
Мышцы конечностей интенсивно развиваются до 5-6 лет и в период полового созревания, причем в первую очередь дифференцируются мышцы кисти и стопы.
Не нашли то, что искали? Воспользуйтесь поиском:
источник
ВОЗРАСТНЫЕ ОСОБЕННОСТИ СКЕЛЕТНОЙ МУСКУЛАТУРЫ
Изменения макро- и микроструктуры скелетных мышц с возрастом.Формирование скелетных мышц происходит на очень ранних этапах развития. На 8-й неделе внутриутробного развития различимы уже все мышцы, а к 10-й неделе развиваются их сухожилия. Связь первичной закладки мышц с соответствующими нервами обнаруживается уже на 2-м месяце развития. Однако двигательные нервные окончания впервые появляются лишь на 4-м месяце внутриутробного развития.
Созревание мышечных волокон связано с увеличением количества миофибрилл, появлением поперечной исчерченности, увеличением числа ядер. Оно осуществляется в разных мышечных волокнах с неодинаковой скоростью. Раньше всего дифференцируются волокна мышц языка, губ, межреберных мышц, мышц спины и диафрагмы. Затем — мышцы верхней конечности и в последнюю очередь — мышцы нижней конечности.
У новорожденных масса мышц составляет 23,3% (у взрослых — 44,2%) от массы всего тела. Сухожильная часть мышцы развита слабо и составляет меньшую, чем у взрослых, часть от всей длины мышцы; фасции и сухожилия широких’ мышц очень тонки, непрочны, легко от них отделяются.
В процессе постнатального развития происходят дальнейшие изменения как макро-, так и микроструктуры скелетных мышц. Созревание различных мышц и даже различных пучков волокон одной и той же мышцы происходит с разной скоростью. Эта скорость определяется функцией, которую выполняет данное анатомическое образование на том или ином возрастном этапе.
Как правило, раньше всего созревают функционально активные мышцы. В целом масса мышц за весь период развития увеличивается примерно на 21%. К 8 годам масса мышц по отношению к массе всего тела становится равной 27,2%, к периоду полового созревания— 32,6%, а в 17—18 лет — 44,2%.
Наиболее интенсивная прибавка массы происходит в период полового созревания. К моменту рождения ребенка наибольшего развития достигают мышцы туловища, головы, верхних конечностей. Их масса составляет около 40% от массы всех мышц (у взрослых — до 30%).
Масса мышц верхних конечностей по отношению к массе мышц всего тела возрастает от рождения до 23—25 лет, когда заканчивается онтогенетическое созревание мышц, всего лишь на 2%. Следовательно, к моменту рождения они уже обладают достаточно большой массой, и дальнейшее увеличение ее шло в полном соответствии с увеличением массы всего тела.
Масса мышц нижних конечностей по отношению к массе тела увеличивается за весь период развития более чем на 16%. В мышцах верхних конечностей особенно резко увеличивается в дошкольном и младшем школьном возрасте масса тех из них, которые вызывают движения пальцев.
Масса мышц-разгибателей увеличивается интенсивнее, чем сгибателей, так как к моменту рождения сгибатели, обусловливающие в период внутриутробного развития характерную позу плода, уже значительно развиты. Разгибатели, обеспечивающие вертикальное положение тела, интенсивно созревают после рождения ребенка.
Мышцы, которые обусловливают большой размах движения, интенсивно растут в длину, а мышцы, функция которых требует сокращений большой силы, увеличиваются в диаметре. Развитие их характеризуется ростом степени перистости.
С возрастом происходит увеличение длины сухожилия. Так, у новорожденного длина сухожилия икроножной мышцы по отношению к длине всей мышцы составляет 0,25, а в 1,5 года — 0,5 (рис. 96). К 12—14 годам отношение длины сухожилия и брюшка мышцы становится таким же, как и у взрослых.
Рост мышц в длину может продолжаться до 23—25 лет. Он осуществляется за счет зоны роста, расположенной на границе мышечной и сухожильной частей. В зоне роста имеется скопление ядер, число которых с возрастом уменьшается, причем особенно значительно после 7 лет. К 15—18 годам зона роста уменьшается в 3 раза. К 15—16 годам заканчивается формирование сарколеммы, когда ее волокна приобретают определенную ориентацию; они направлены перпендикулярно к продольной оси мышечного волокна. Развитие в постнатальном онтогенезе соединительной ткани мышц характеризуется уменьшением числа клеток, приходящихся на единицу площади, и увеличением числа волокон. Так, к 3—4 годам число клеток соединительной ткани, приходящихся на единицу площади, составляет 25—30 (у новорожденного — 50—60). Даже в 13—15 лет число клеток все еще в 2 раза больше, чем у взрослых.
Диаметр мышечных волокон увеличивается до 35 лет. Изменение диаметра каждого отдельного мышечного волокна и общего числа их вызывает нарастание поперечника мышц в процессе роста и развития организма. Разница в диаметре волокон одной и той же мышцы отмечается уже в эмбриональном периоде развития. Так, в двуглавой мышце плеча на 5—6 месяце внутриутробного развития встречаются волокна диаметром и 2,5, и 12,5 мкм. Но осо-бенно большой эта разница становится после рождения. Например, толщина волокон большой ягодичной мышцы и прямой мышцы глаза у новорожденных одинакова, а у взрослых диаметр волокон одной мышцы больше диаметра другой в 5 раз. Диаметр одних волокон увеличивается в 5—9 раз (двуглавая мышца плеча), других— в 4—5 раз (сгибатель пальцев и кисти).
Наиболее интенсивный рост волокон отмечается в период полового созревания.
Так, в первые 6 месяцев жизни диаметр волокон двуглавой мышцы плеча составляет 17—19 мкм, в 2—3 года— 20—22 мкм, в 9—12 лет—20—25 мкм, а у взрослых — 41 — 58 мкм.
С возрастом растет число миофибрилл за счет их продольного расщепления. У новорожденного в каждом мышечном волокне содержится 50—120 миофибрилл, в 1,5 года их число становится в 2 раза большим, в 3—4 года оно увеличивается в 5—6 раз, к 7 годам— в 15—20 раз (Л. К. Семенова).
Химический состав мышц с возрастом также меняется. Мышцы детей содержат больше воды, они богаты нуклеопротеидами. По мере роста происходит неуклонное нарастание актомиозина и АТФ, креатинфосфорной кислоты, мышечного гемоглобина (миоглобина). В связи с тем что миоглобин является источником кислорода, увеличение его количества способствует совершенствованию сократительной функции мышцы.
Развитие двигательных нервных окончаний происходит особенно интенсивно в первые месяцы после рождения. Но только к 11 — 13 годам полностью заканчивается их структурное оформление.
Не нашли то, что искали? Воспользуйтесь поиском:
источник
Строение, иннервация и функции скелетных мышц
Строение мышцы. Мышца состоит из пучков поперечнополосатых мышечных волокон, соединенных рыхлой соединительной тканью в пучки первого порядка. Они, в свою очередь, объединяются в пучки второго порядка и т. д. В итоге мышечные пучки всех порядков объединяются соединительной оболочкой, образуя мышечное брюшко. Соединительнотканные прослойки, имеющиеся между мышечными пучками по концам брюшка, переходят в сухожильную часть мышцы, крепящейся к кости. Во время сокращения происходит укорочение мышечного брюшка и сближение ее концов. При этом сократившаяся мышца с помощью сухожилия тянет за собой кость, которая выполняет роль рычага. Так совершаются разнообразные движения
Функции мышц. Мышцы — это органы тела, состоящие из мышечной ткани, способной сокращаться под влиянием нервных импульсов. Они являются активным элементом опорно-двигательной системы, так как обеспечивают разнообразные движения при перемещении человека в пространстве, сохранение равновесия, дыхательные движения, сокращения стенок внутренних органов, голосообразование и др.
Иннервация мышц.Скелетные мышцы получают двигательную, чувствительную и трофическую (вегетативную) иннервацию.
Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы — от двигательных нейронов определенных черепных нервов. При этом к каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется одним мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже сотни мышечных волокон получают двигательную иннервацию от одного мотонейрона, посредством разветвления его аксона.
Двигательное нервное волокно, подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обуславливая выход ионов кальция и начало процесса сокращения мышечного волокна.
Чувствительная (афферентная) иннервация скелетных мышц осуществляется псевдоуниполярными нейронами спинальных ганглиев, посредством разнообразных рецепторных окончаний дендритов этих клеток.
Строение, иннервация и функции висцеральных мышц.
Висцеральные мышцы, действующие непроизвольно и лишенные поперечных полос, располагаются прежде всего в стенках пищеварительной трубки. Они отвечают за перистальтические движения, которые проталкивают пищу в пищеварительном тракте.
Висцеральная гладкая мышца имеет двойную иннервацию— симпатическую и парасимпатическую, функция которой заключается в изменении деятельности гладкой мышцы. Раздражение одного из вегетативных нервов обычно увеличивает активность гладкой мышцы, стимуляция другого — уменьшает.
Учение о костях и их соединениях.
Строение костей
Скелет как опора несет большой груз: в среднем 60-70 кг (масса тела взрослого человека). Поэтому кости должны быть прочными. Кости выдерживают растяжение почти также, как чугун, а по сопротивлению на сжатие они вдвое превосходят гранит.
Мягкие части кости не делают ее менее прочной. Клетки костной ткани живут как бы одной семьей, соединяясь друг с другом отростками, как мостиками. Кровеносные сосуды, пронизывая кость и доставляя костным клеткам питательные вещества и кислород, не снижают надежной твердости кости.
В трубчатых костях повышению их прочности служат и различия в структуре по направлению от центра к концам. Трубчатая кость в центре отличается большей твердостью и меньшей эластичностью, чем на концах. По направлению к суставной поверхности структура трубчатой кости меняется от компактной к плотной. Такое изменение строения обеспечивает главную передачу напряжения от кости через хрящ на поверхность сустава.
Снаружи кость одета надкостницей, которую пронизывают кровеносные сосуды, питающие кость. В надкостнице имеется много чувствительных нервных окончаний, сама же кость нечувствительна.
Полость трубчатых костей заполнена красным костным мозгом, который в течение жизни заменяется желтым (жировой тканью).
Формы костей.
Форма костей
Кости отличаются друг от друга по форме и строению. Выделяют кости трубчатые, плоские, смешанные и воздухоносные. Среди трубчатых костей различают длинные (плечевая, бедренная, кости предплечья, голени) и короткие (кости пясти, плюсны, фаланги пальцев). Губчатые кости состоят из губчатого вещества, покрытого тонким слоем компактного вещества. Они имеют форму неправильного куба или многогранника и располагаются в местах, где большая нагрузка сочетается с подвижностью (например, надколенник).
Плоские кости участвуют в образовании полостей, поясов конечностей и выполняют функцию защиты (кости крыши черепа, грудина).
Смешанные кости имеют сложную форму и состоят из нескольких частей, имеющих разное происхождение. К смешанным костям относятся позвонки, кости основания черепа.
Воздухоносные кости имеют в своем теле полость, выстланную слизистой оболочкой и заполненную воздухом. Таковы, например, некоторые части черепа: лобная, клиновидная, верхняя челюсть и некоторые другие.
Форма и рельеф костей зависят от характера прикрепления к ним мышц. Если мышца прикрепляется к кости с помощью сухожилия, то в этом месте формируется бугор, отросток или гребень. Если же мышца непосредственно сочетается с надкостницей, то образуется углубление.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Для студентов недели бывают четные, нечетные и зачетные. 9537 —  | 7473 —
| 7473 —  или читать все.
или читать все.
85.95.179.227 © studopedia.ru Не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования. Есть нарушение авторского права? Напишите нам | Обратная связь.
Отключите adBlock!
и обновите страницу (F5)
очень нужно
источник
Иннервация скелетной мышцы
Рассмотрена иннервация скелетной мышцы. Описан состав и строение нейрона, а также функции нейрона. Рассмотрены функции двигательных и чувствительных нейронов. Описан состав периферического нерва. Введено понятие двигательной единицы и синапса.
Иннервация скелетной мышцы
Иннервация (от in — в, внутри и nervus — нервы) — снабжение органов и тканей нервами, что обеспечивает их связь с центральной нервной системой (ЦНС).
Иннервация скелетной мышцы — это наличие нервных волокон, которые передают импульсы из ЦНС к мышце и от мышцы в ЦНС.
Денервация скелетной мышцы — нарушение передачи импульсов из ЦНС к мышце или от мышцы в ЦНС. Денервация мышечных волокон скелетной мышцы возможна из-за перерезки или повреждения нерва. Денервация мышечных волокон происходит также по мере старения из-за уменьшения количества нервных волокон, иннервирующих мышцу.
Теперь рассмотрим этот вопрос подробнее. Давайте сначала разберемся, что представляет собой нейрон.
Нейрон – это структурная единица нашей нервной системы, главная функция которого – передача информации от одного участка тела другому. Чтобы передать информацию нейрон возбуждается, затем вырабатывает нервный импульс. Нейроны также участвуют в обработке и хранении информации. Но этого вопроса мы касаться не будем.
Нейрон – это высокоспециализированная клетка. Он состоит из тела и длинного отростка – аксона (рис. 1.). 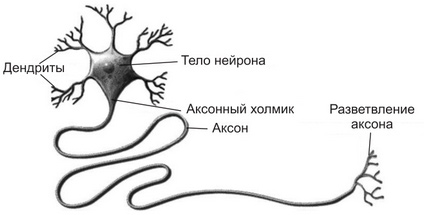 Длина аксона может достигать одного метра и более. На теле есть много мелких отростков – дендритов. Через эти отростки нейрон получает информацию от других нейронов и передает ее через аксон или другому нейрону, или органу, к которому он подходит (в нашем случае – мышце).
Длина аксона может достигать одного метра и более. На теле есть много мелких отростков – дендритов. Через эти отростки нейрон получает информацию от других нейронов и передает ее через аксон или другому нейрону, или органу, к которому он подходит (в нашем случае – мышце).
Двигательные нейроны (мотонейроны) несут информацию от ЦНС мышце. Тела мотонейронов расположены в передних рогах спинного мозга. Аксоны мотонейронов идут к мышцам в составе периферического нерва.
Чувствительные нейроны передают информацию о состоянии мышцы в ЦНС. Тела чувствительных нейронов расположены в задних рогах спинного мозга.
Более подробно строение мышцы рассмотрено в моих книгах «Гипертрофия скелетных мышц человека» и «Биомеханика мышц«
Большинство периферических нервов являются смешанными. Смешанный нерв в своем составе содержит как двигательные, так и чувствительные волокна. Структура периферического нерва похожа на структуру мышцы.
Когда мы рассматривали строение мышцы, то указывалось, что периферические нервы имеют каналы в перимизии. Через эти каналы периферические нервы подходят к мышечным волокнам.
Один аксон иннервирует (то есть пускает веточки) достаточно много мышечных волокон. Иногда на один аксон приходится более 2000 мышечных волокон. Такая система, состоящая из одного нейрона и мышечных волокон, которые он иннервирует, называется двигательной единицей (ДЕ). Это понятие ввел в физиологию нобелевский лауреат Чарльз Скотт Шеррингтон в начале XX века. Особенности состава и функционирования ДЕ мы рассмотрим позднее.
По двигательным волокнам к мышце поступают импульсы из центральной нервной системы (ЦНС) в результате которых мышца развивает возбуждение (сокращается). Чувствительные волокна несут в ЦНС информацию о различных показателях активности мышцы (длине мышцы, скорости ее сокращения, степени напряжения). Если бы ЦНС не могла получать эту информацию, управление напряжением мышцы было бы невозможно. Точно так же было бы невозможно управление нашими движениями.
Место, где аксон мотонейрона соединяется с мышечным волокном называется синапсом. Через синапс к мышечному волокну поступают сигналы, которые вызывают его возбуждение.
Иннервация скелетных мышц пожилых людей ухудшается. Ученые находят, что с возрастом уменьшается количество мотонейронов, иннервирующих скелетную мышцу. Это является одним из факторов, обусловливающих возрастное уменьшение массы мышц и их силы — саркопению.
- Ванек, Ю. Спортивная анатомия.– М.: Академия, 2008.- 304 с.
- Мак-Комас, А.Дж. Скелетные мышцы.– Киев: Олимпийская литература, 2001.– 407 с.
- Самсонова, А.В. Гипертрофия скелетных мышц человека. – СПб: Кинетика, 2018. – 159 с.
источник
Возрастные особенности функции скелетных мышц
Морфологический субстрат сократительного акта формируется на ранних стадиях онтогенеза. Скелетные мышцы уже к семи неделям внутриутробного развития способны сокращаться, а к 12 неделям возникают элементарные рефлекторные движения в ответ на механические раздражения. В 12 недель плод может сжимать кисть в кулак.
Сокращение в эмбриональный период развития носит тонический характер: оно медленно нарастает и затем длительно удерживается на одном уровне. Такой характер реакции обусловливается содержанием в мышце большого количества волокон типа «красных».
Процесс возбуждения мышечного волокна, предшествующий его сокращению, приобретает характеристики, свойственные мышцам взрослых, не сразу. У детей мембранный потенциал и ПД отличаются по своим характеристикам от таковых у взрослых. Увеличение содержания ионов внутриклеточного калия происходит до 9 лет. Поскольку ионы К определяют величину мембранного потенциала покоя, значение ее в течение первых 9 лет жизни у детей меньше, чем у взрослых.
Полное функциональное созревание мышц происходит на поздних этапах развития. Электровозбудимость мышц детей в эмбриональный период и в первые месяцы постнатального развития меньше, чем у взрослых, т. е. порог раздражения более высокий.
Чем меньше возраст детей, тем меньше величина лабильности их мышечных волокон. Отсюда следует, что длительность процессов реполяризации с возрастом увеличивается.
Созревание волокон одной и той же мышцы происходит неодновременно. Часть волокон имеет лабильность, характерную для мышц взрослых уже в возрасте 10—13 лет, а лабильность других волокон продолжает увеличиваться вплоть до 14—15 лет.
Прямые определения длительности ПД в отдельных волокнах мышцы показали, что величина его в возрасте 14 лет составляет 5,7 мс против 2 мс у взрослых. С возрастом увеличивается мышечная сила за счет роста диаметра мышечных волокон и их числа.
Для разных мышц наибольшая величина их силы отмечается в разном возрасте. Большинство мышц верхней конечности и шеи достигает наибольшей силы к 20—30 годам. В то же время сила мышц, разгибающих туловище, становится максимальной к 16 годам. Разница между силами мышц-сгибателей и мышц-разгибателей увеличивается с возрастом.
Характеристикой функционального созревания мышц служит мышечная выносливость. В дошкольном и младшем школьном возрасте происходит наибольшее по сравнению с другими возрастами увеличение выносливости. Однако даже в 16—19 лет ее величина составляет лишь 85% от выносливости взрослого.
Изучение характера электрической активности мышцы при произвольных максимальных ее напряжениях позволило считать, что меньшая способность детей сохранять максимальную величину статического усилия зависит от быстро развивающегося у них процесса асинхронизации работы мышечных волокон. Чем меньше возраст, тем меньшее время может сохраняться одновременное возбуждение волокон мышцы и тем быстрее в ней наступает утомление.
Морфологическое и функциональное созревание опорно-двигательного аппарата зависит от очень многих факторов: наследственных, условий жизни, питания, двигательной активности. Правильно организованные занятия физической культурой и спортом способствуют формированию как костной, так и мышечной системы. Под влиянием этих занятий увеличивается диаметр мышечных волокон, растет их число, более совершенными становятся координационные отношения между мышцами-антагонистами. Однако следует помнить, что мышечная деятельность вызывает у детей по сравнению со взрослыми значительно большие изменения в деятельности всех их органов, что требует дифференцированного подхода к организации всякого рода занятий физкультурой с детьми различного возраста.
Не нашли то, что искали? Воспользуйтесь поиском:
источник
ИННЕРВАЦИЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Скелетные мышцы обладают эфферентной (двигательной) и афферентной (чувствительной) иннервацией.
Эфферентная иннервацияскелетных мышц обеспечивается нервными волокнами (аксонами а-мотонейронов), образующими на мышечных волокнах специализированные нервно-мышечные окончания (нервно-мышечные синапсы, или моторные бляшки), которые осуществляют передачу возбуждения с нервного волокна на мышечное. Один мотонейрон может иннервировать различное количество мышечных волокон.
Двигательная единица(ДЕ) представляет собой совокупность мотонейрона и иннервируемых им мышечных волокон. Количество мышечных волокон, входящих в одну ДЕ, минимально в мелких мышцах, осуществляющих точные и тонкие движения. В глазных мышцах человека ДЕ включает от 2-6 до 13-20 мышечных волокон, в мышцах кисти – от 10-25 до 100-300, в мышцах туловища – 1500-2000. Управление мышечной активностью обеспечивается как изменением частоты активации ДЕ, так и вовлечением различного их числа в процесс сокращения.
Мышечные волокна, образующие одну ДЕ, обладают одинаковыми механическими свойствами, гистохимическими характеристиками и относятся к одному типу, однако рассеяны по обширной территории мышцы.
Афферентная иннервацияскелетных мышц обеспечивается нервно-мышечными веретенами — рецепторами растяжения волокон поперечнополосатых мышц, которые представляют собой сложные инкапсулированные нервные окончания, состоящие из веточек нервных волокон, оплетающих особые тонкие (интрафузальные) мышечные волокна, заключенные в тончайшую соединительнотканную капсулу. Остальные мышечные волокна назваются экстрафузальными.
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Сердечная мышечная тканьвстречается только в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Ее клетки (сердечные миоциты, или кардиомиоциты) составляют лишь 30-40% общего числа клеток сердца, но образуют 70-90% его массы. Основным функциональным свойством сердечной мышечной ткани служит способность к спонтанным ритмическим сокращениям, на активность которых влияют гормоны и нервная система (симпатическая и парасимпатическая).
 А А |
 Б Б |
Рис. 7.12. На электронной микрофотографии изображен продольный срез миокарда А(х 198), Б(х 640). Ядра в миоцитах расположены центрально, синтициальное строение миокарда (образуется симпласт) – миоциты соединяются между собой с помощью вставочных дисков (D). Между миоцитами расположены прослойки соединительной ткани, в которой много капилляров (С).
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ СЕРДЕЧНОЙ
МЫШЕЧНОЙ ТКАНИ
Сердечная мышечная тканьобразована клетками — кардиомиоцитами(сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон (рис. 7.12).
Кардиомиоциты — цилиндрические или ветвящиеся клетки, более крупные в желудочках, где их длина составляет 100-150 мкм, а диаметр — 10-20 мкм. В предсердиях они обычно имеют неправильную форму и меньшие размеры (длина — 40-70 мкм, диаметр — 5-6 мкм). Кардиомиоциты содержат одно или два ядра и саркоплазму, покрыты сарколеммой, которая снаружи окружена базальной мембраной.
Саркоплазма кардиомиоцитов содержит органеллы и включения, которые образуют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппаратсильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых), которых он занимает до 50-70% объема клетки. Фагоструктура сходна с таковой поперечно-полосатой мышечной ткани. Механизм мышечного сокращения этих видов мышечной ткани также сходны.

Рис. 7.13. Саркотубулярная система кардиомиоцита. Саркотубулярная система представлена саркоплазматической сетью (СПС), не образующей терминальных цистерн, и широкими Т-трубочками (Т-ТР).
Аппарат передачи возбуждения (саркотубулярная система)в кардиомиоцитах в целом сходен с таковым в скелетных мышечных волоконах, однако он обладает рядом особенностей (рис. 7.13).
Саркоплазматическая сеть развита слабее, чем в скелетном мышечном волокне, менее активно накапливает Са 2+ , не образует терминальных цистерн. Во время расслабления она выделяет ионы Са 2+ в саркоплазму с низкой скоростью, что обеспечивает автоматизм кардиомиоцитов (рис. 7.14).
 А А |
 Б Б |
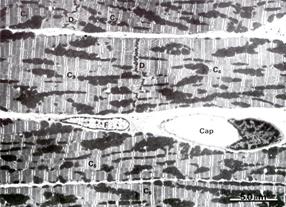
Рис. 7.14. На электронной микрофотографии изображен: А (х5.000) – участок миокарда состоящий из 6 миоцитов (С1-С6). Эти клетки соединяются между собой вставочным диском D. В поле зрения в миоцитах нет ядер. Между миоцитами – рыхлая соединительная ткань, в которой много капилляров (Сар), фибробластов (F); Б (х38.000) ультраструктура саркомера миокарда схожа с таковой поперечно-полосатой мышечной тканью. Обратить внимание на строение А и J дисков СПР (SR), тубулярной системы (Т), расположение митохондрий. В миоците много липидных капель (L) и гранул гликогена (G).
Поперечные (Т-) трубочки — широкие, содержат компоненты базальной мембраны, вместе с элементами саркоплазматической сети образуют диады (включают одну
Т-трубочку и одну цистерну сети), которые располагаются в области Z-линий (рис. 7.14).
Опорный аппарат кардиомиоцитовпредставлен элементами цитоскелета, обеспечивающими упорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также базальной мембраной и сарколеммой. Его структурная и биохимическая организация сходна с таковой в волокнах скелетной мышечной ткани. В кардиомиоцитах его особенностью служит то, что элементы цитоскелета связаны с особыми межклеточными соединениями – вставочными дисками.
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих функциональное волокно сердечной мышечной ткани (рис. 7.15).
Энергетический аппарат кардиомиоцитовпредставлен митохондриями и включениями, расщепление которых обеспечивает получение энергии. Митохондрии кардиомиоцитов обладают также свойством накапливать кальций в высоких концентрациях.

Рис. 7.15. Ультраструктурная организация области вставочного диска кардиомиоцитов. В поперечных участках вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом (Д) актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска в участке полоски слипания (ПС). На сарколемме продольных участков вставочного диска располагаются щелевые соединения (ЩС). БМ – базальная мембрана, СЛ – сарколемма, МТХ – митохондрия. Обозначения компонентов саркомера СМ) – те же, что на рис. 4.
Синтетический аппарат кардиомиоцитов Синтетический аппарат включает свободные рибосомы и полисомы, цистерны грЭПС. и элементы комплекса Гольджи, которые располагаются в саркоплазме преимущественно у полюсов ядра.
Лизосомальный аппарат (аппарат внутриклеточного переваривания) кардиомиоцитовхорошо развит, что отражает высокую скорость обновления их структурных компонентов. Он включает эндосомы, лизосомы и остаточные тельца. Собственно лизосомы располагаются у полюсов ядра и занимают до 10% объема саркоплазмы. Остаточные тельца в виде липофусциновых гранул очень многочисленны (при старении могут составлять до 20% сухой массы миокарда).
Типы кардиомиоцитов
Сердечная мышечная ткань содержит кардиомиоциты трех основных типов: 1) сократительные (рабочие); 2) проводящие 3) секреторные (эндокринные).
1) сократительные (рабочие) кардиомиоциты образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим большую часть их саркоплазмы;
2) проводящие кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они образуют узлы и пучки проводящей системы сердца и разделяются на несколько подтипов. Характеризуются слабым развитием сократительного аппарата, светлой саркоплазмой и крупными ядрами
3) секреторные (эндокринные) кардиомиоцитырасполагаются в предсердиях (в особенности, правом) и характеризуются отростчатой формой и слабым развитием сократительного аппарата. В их саркоплазме вблизи полюсов ядра находятся окруженные мембраной плотные гранулы диаметром 200-300 нм, содержащие гормон – предсердный натриуретический фактор (пептид) — ПНФ (ПНП). Этот гормон вызывает усиленную потерю натрия и воды с мочой (натриурез и диурез), расширение сосудов, снижение артериального давления, угнетение секреции альдостерона, кортизола и вазопрессина.
Кровоснабжение сердечной мышечной тканичрезвычайно обильно: по уровню кровоснабжения (мл/мин/100 г массы) миокард уступает лишь почке и превосходит другие органы, включая головной мозг.
Иннервация сердечной мышечной тканиосуществляется волокнами симпатического и парасимпатического отделов вегетативной нервной системы.
Регенерация сердечной мышечной тканиу взрослого человека может осуществляться только на внутриклеточном уровне путем обновления структурных компонентов кардиомиоцитов, поскольку их способность к пролиферации утрачивается, по-видимому, еще в раннем детстве.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань очень широко распространена в организме: она входит в состав стенки полых (трубчатых) внутренних органов — бронхов, желудка, кишки, матки, маточных труб, мочеточников, мочевого пузыря (висцеральная гладкая мышечная ткань), а также сосудов (васкулярная гладкая мышечная ткань). Васкулярная гладкая мышечная ткань отличается от висцеральной некоторыми структурными, биохимическими и функциональными особенностями, чувствительностью к действию ряда гормонов, нейромедиаторов и фармакологических препаратов. Гладкая мышечная ткань встречается также в коже, где она образует мышцы, поднимающие волос, а также в капсулах и трабекулах некоторых органов (селезенка, яичко) (рис. 7.16).

Рис. 7.16. На микрофотографии (х480)изображен продольный срез гладкой мышечной ткани. Мышечная клетка имеет веретенообразную форму, в центре расположено ядро. Концы клеток вклиниваются между срединными частями соседних клеток, благодаря чему создается связь клеток в пласте. Между гладкомышечными клетками располагается соединительная ткань.
Движения, осуществляемые гладкой мышечной тканью, – сравнительно медленные и продолжительные, она обеспечивает также длительные тонические сокращения. Ее сокращения вызывают изменения величины просвета трубчатых органов и лежат в основе их перистальтики. Благодаря сократительной активности этой ткани обеспечивается деятельность органов пищеварительного тракта, регуляция дыхания, крово- и лимфотока, выделение мочи, транспорт половых клеток и др.
Последнее изменение этой страницы: 2016-04-21; Нарушение авторского права страницы
источник