Мир психологии
психология для всех и каждого
Сердечно-сосудистая система. Часть 4.

В этой части речь идет о свойствах сердечной мышцы: возбудимости и возбуждении сердечной мышцы, об особенностях рефрактерного периода сердечной мышцы, об автоматии, о механизме автоматии, о скорости проведения возбуждения в сердце, о сократимости сердечной мышцы.
Свойства сердечной мышцы.
Сердечная мышца обладает возбудимостью, способностью генерировать потенциал действия, проводить возбуждение, сокращаться и др. Одно из важнейших свойств сердечной мышцы — автоматия.
Возбудимость и возбуждение сердечной мышцы.
Возбудимость сердечной мышцы меньше, чем скелетной: она обладает более высоким порогом раздражения, более длительным латентным и рефрактерным периодами и больше величиной хронаксии.
Величина мембранного потенциала значительно отличается в разных участках сердца. В мышечных волокнах предсердий она составляет 80-90 мв, в волокнах желудочков и пучка Гисса 90 мв, а в волокнах Пуркинье — 96 мв, т.е. величина мембранного потенциала различных волокон сердечной мышцы больше величины мембранного потенциала скелетной мышцы. Для синоатриального и атриовентрикулярного узла характерна меньшая величина мембранного потенциала — 50-65 мв.
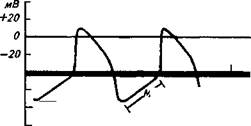
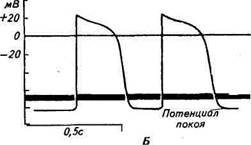
При возбуждении возникает потенциал действия скелетной мышцы. В разных структурах сердца различны его величина и форма. В среднем амплитуда потенциала действия составляет 100-120 мв.
Форма потенциала действия мышечных волокон желудочков и предсердий имеет значительные отличия от потенциала действия скелетной мышцы или нерва.
В потенциале действия сердечной мышцы различают фазу быстрой деполяризации, во время которой после достижения нулевого уровня имеет место реверсия потенциала. Фаза быстрой деполяризации сменяется фазой длительной реполяризации, в которой различают быструю реполяризацию, сменяющуюся длительно идущей фазой медленной реполяризации, или плато, переходящей в фазу конечной быстрой реполяризации. Затем наступает завершающий момент — фаза диастолического расслабления. Последняя отделяет одно сокращение от другого.
Длительность потенциала действия мышечных волокон сердца значительно больше, чем волокон скелетной мышцы. В среднем она равна 0,3 сек при 70 сокращениях сердца в минуту и изменяется с изменением частоты сердечных сокращений. При уменьшении частоты сокращений сердца длительность потенциала действия увеличивается, а при увеличении частоты сокращений она уменьшается.
Иной характер потенциала действия в синоатриальном и атриовентрикулярном узлах проводящей системы сердца. Его величина невелика (50-65 мв), в нем отсутствуют плато и фаза диастолического расслабления. В потенциале действия синусного узла выделяют две основные фазы: фазу медленной деполяризации и фазу медленной реполяризации. Характерной особенностью этого потенциала является наличие фазы спонтанной деполяризации, сменяющей фазу реполяризации. При этом в синоатриальном узле проводящей системы сердца самопроизвольно наступает деполяризация, которая достигает критического уровня и приводит к возникновению одного потенциала действия за другим.
Возникновение потенциала действия сердечной мышцы связано с изменением проницаемости мембраны. Во время диастолы увеличивается проницаемость синоатриального узла к ионам натрия и уменьшается по отношению к ионам калия. При этом происходит деполяризация мембраны. В фазу реполяризации увеличивается проницаемость мембраны по отношению к ионам калия, в результате чего восстанавливается заряд мембраны. В возникновении возбуждения в сердечной мышцы принимают участие ионы кальция. Перемещение ионов натрия и калия происходит не только пассивно вследствие разности их концентраций, но и с участием активных механизмов (большое значение имеет фермент АТФаза).
Особенности рефрактерного периода сердечной мышцы.
Сердечная мышца обладает длительным рефрактерным периодом. Абсолютный рефрактерный период длится почти весь период сокращения сердца, он соответствует систоле. При 70 сокращениях сердца в минуту длительность его равна 0,27 сек. В связи с этим раздражение, нанесенное на сердце в момент систолы, остается без ответа. Сердечная мышца отвечает на раздражение только в момент окончания систолы или в период диастолы. А поэтому она отвечает только на одиночное раздражение и в обычных условиях деятельности сердечная мышца не способна в ответ на ритмическое раздражение развивать длительное непрерывное сокращение, называемое тетанусом.
Абсолютный рефрактерный период сменяется относительным, соответствующим концу систолы, и длится 0,03 сек. Затем следует очень короткий период повышенной возбудимости — фаза экзальтации (или супернормальности), во время которой сердечная мышца может отвечать возбуждением и на подпороговое раздражение. После этого восстанавливается исходный уровень возбудимости сердечной мышцы.
Автоматия.
Автоматией называют способность клетки, ткани, органа возбуждаться без участия внешнего стимула, под влиянием импульсов, возникающих в них самих.
Показателем автоматии сердечной мышцы может быть тот факт, что изолированное сердце лягушки, удаленное из организм и помещенное в физиологический раствор, может в течение длительного времени ритмически сокращаться.
Различные отделы сердца обладают разной способностью к автоматии. Самой высокой автоматией обладает синоатриальный узел. От его активности зависит частота сердечных сокращений, в связи с чем его называют ведущим узлом сердца или водителем ритма.
Меньшей способностью к автоматии обладает атриовентрикулярный узел и еще меньшей — пучок Гисса. Способность к ритмической активности разных отделов проводящей системы сердца можно четко наблюдать в опытах Станниуса с наложением лигатур — перевязок. В опыте на лягушке с помощью лигатуры отделяется часть предсердия вместе с синоатриальным узлом от остальной части сердца. После этого все сердце перестает сокращаться, а отделенный участок предсердия продолжает сокращаться в том же ритме, что и до наложения лигатуры. Это говорит о том, что синоатриальный узел является ведущим, от него зависит частота сердечных сокращений.
Через некоторое время (20-30 мин) после наложения лигатуры на сердце лягушки проявляется автоматия атриовентрикулярного узла: сердце начинает сокращаться, но в боле редком ритме, чем до наложения лигатуры, причем предсердия и желудочки сокращаются одновременно.
Если на сердце теплокровного животного создать блок между атриовентрикулярным узлом и пучком Гисса, то верхушка сердца будет сокращаться в еще более редком ритме, который зависит от автоматии пучка Гисса или волокон Пуркинье.
Из изложенного можно сделать вывод, что способность сердца к автоматии уменьшается от венозного конца сердца к артериальному. Эта особенность была отмечена Гаскеллом и названа им законом градиента сердца.
В нормальных условиях жизнедеятельности организма проявляется автоматия только синоатриального узла и ему подчинены все другие отделы сердца, из автоматия подавляется водителем ритма.
Механизм автоматии.
Ритмической активностью в сердце обладают элементы атипической ткани и мышечные клетки. Способность к автоматии индивидуальна и закладывается в самые ранние периоды эмбрионального развития сердца. Было показано, что отдельные мышечные волокна сердца могут сокращаться в разном ритме, но, как только они объединяются морфологически, наиболее быстро сокращающаяся клетка берет на себя функцию водителя ритма.
В основе ритмической автоматии лежит способность клеток проводящей системы сердца к спонтанной деполяризации, к спонтанному изменению мембранного потенциала, которое наступает в конце фазы реполяризации и при достижении критического уровня приводит к возникновению нового потенциала действия и, соответственно, нового сокращения. Чем быстрее возникает деполяризация, тем больше частота сердечных сокращений.
В основе спонтанной диастолической деполяризации лежат до конца еще не изученные ионные механизмы проницаемости мембраны клеток — водителей ритма по отношению к ионам натрия и калия.
Скорость проведения возбуждения в сердце.
Сокращение мышечных волокон сердца вызывается импульсами, автоматически возникающими в синоатриальном узле. Возникший здесь потенциал действия распространяется на мышцы предсердий, затем к атриовентрикулярному узлу, от него — к пучку Гисса и далее по волокнам Пуркинье переходит на миокард правого и левого желудочков.
В различных участках сердца скорость проведения возбуждения неодинакова. Она зависит от количества десмосом, которые обладают малым сопротивлением (оно в 100 раз меньше, чем в сарколемме) и тем способствуют большой скорости проведения возбуждения. Десмосом в синоатриальном узле мало, и поэтому скорость проведения возбуждения в нем невелика — 0,05 м/сек. От синоатриального узла потенциал действия распространяется по волокнам правого и левого предсердий к перегородке между ними. Скорость проведения возбуждения по мышцам предсердий 1 м/сек. Оба предсердия оказываются охваченными возбуждением через 0,12 сек.
От предсердий возбуждение переходит к атриовентрикулярному узлу. Здесь оно возникает не сразу и происходит некоторая задержка в проведении возбуждения. Она имеет важное функциональное значение, так как способствует определенной последовательности сокращений различных отделов сердца. Желудочки сокращаются только после того, как закончилось сокращение предсердий. Относительно механизмов атриовентрикулярной задержки существует ряд мнений, основанных на морфологических и функциональных особенностях этого отдела сердца. С помощью микроэлектродной техники установлено, что в области атриовентрикулярного узла имеется синапс, в котором самостоятельно развивается возбуждение. Как любой синапс, синапс в атриовентрикулярном узле обладает более низкой возбудимостью, односторонним и замедленным проведением возбуждения. Вследствие небольшой возбудимости синапса импульс, приходящий к нему от предсердий, оказывается подпороговым. Необходима суммация подпороговых импульсов, для того, чтобы возникло распространяющееся возбуждение. Время суммации возбуждения (суммация подпороговой деполяризации) составляет время атриовентрикулярной задержки.
По структурам атриовентрикулярного узла возбуждение проводится со скоростью 0,08 м/сек, пучка Гисса 0 1,5 м/сек. Наибольшей скоростью проведения возбуждения обладают волокна Пуркинье — 4-5 м/сек, так как в них содержится большое количество десмосом. В мышцах желудочков скорость проведения снова уменьшается, она составляет 0,5-0,8 м/сек.
Сократимость сердечной мышцы.
Сократимостью обладают мышечные волокна сердца — миофибриллы. Сигналом к их сократительной деятельности является возникновение в них возбуждения. Возбуждение, возникнув в сарколемме мышечного волокна, распространяется по системе саркоплазматического ретикулюма внутрь волокна и вызывает его сокращение. В основе сокращения мышечных волокон сердца лежит тот же механизм, что и в основе сокращения скелетных мышц, — скольжение нитей актина и миозина.
Сердечная мышца отвечает на раздражение в соответствии с правилом «все или ничего», т.е. при достижении пороговой величины раздражения сердце отвечает максимальным сокращением и с увеличением силы раздражения величина ответа не изменяется. В этом характерная особенность ее сокращения. Правда, величина максимального ответа может быть различной и зависит от функционального состояния мышцы.
Величина сокращения сердечной мышцы зависит от первоначальной длины ее волокон. Эта зависимость выражается «законом сердца» Старлинга: сила сокращения тем больше, чем больше первоначальное растяжение мышечных волокон сердца. При увеличении притока крови к сердцу увеличивается растяжение его волокон и увеличивается сила сердечных — сердце больше выбрасывает крови за одно сокращение. Данное свойство имеет большое значение в приспособлении сердца к различным условиям деятельности при выполнении физической или спортивной нагрузки, изменении положения тела и т.д.
источник
Сердечная мышца человека











- Физиология

- История физиологии
- Методы физиологии



Физиологические свойства сердечной мышцы
Кровь может выполнять свои многочисленные функции, только находясь в постоянном движении. Обеспечение движения крови является главной функцией сердца и сосудов, формирующих кровеносную систему. Сердечно-сосудистая система совместно с кровью участвует также в транспорте веществ, терморегуляции, реализации иммунных реакций и гуморальной регуляции функций организма. Движущая сила кровотока создастся за счет работы сердца, которое выполняет функцию насоса.
Способность сердца сокращаться в течение всей жизни без остановки обусловлена рядом специфических физических и физиологических свойств сердечной мышцы. Сердечная мышца уникальным образом сочетает в себе качества скелетной и гладкой мускулатуры. Так же как и скелетные мышцы, миокард способен интенсивно работать и быстро сокращаться. Так же как и гладкие мышцы, он практически неутомим и не зависит от волевого усилия человека.

Физические свойства
Растяжимость — способность увеличивать длину без нарушения структуры под влиянием растягивающей силы. Такой силой является кровь, наполняющая полости сердца во время диастолы. От степени растяжения мышечных волокон сердца в диастолу зависит сила их сокращения в систолу.
Эластичность — способность восстанавливать исходное положение после прекращения действия деформирующей силы. Эластичность сердечной мышцы является полной, т.е. она полностью восстанавливает исходные показатели.
Способность развивать силу в процессе сокращения мышцы.
Физиологические свойства
Сокращения сердца происходят вследствие периодически возникающих процессов возбуждения в сердечной мышце, которая обладает рядом физиологических свойств: автоматизмом, возбудимостью, проводимостью, сократимостью.
Способность сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом, носит название автоматизм.
В сердце различают сократительную мускулатуру, представленную поперечно-полосатой мышцей, и атипическую, или специальную ткань, в которой возникает и проводится возбуждение. Атипическая мышечная ткань содержит малое количество миофибрилл, много саркоплазмы и не способна к сокращению. Она представлена скоплениями в определенных участках миокарда, которые образуют проводящую систему сердца, состоящую из синоатриального узла, располагающегося на задней стенке правого предсердия у места впадения полых вен; атриовентрикулярного, или предсердно-желудочкового узла, находящегося в правом предсердии вблизи перегородки между предсердиями и желудочками; предсердно-желудочкового пучка (пучка Гиса), отходящего от атриовентрикулярного узла одним стволом. Пучок Гиса, пройдя через перегородку между предсердиями и желудочками, разветвляется на две ножки, идущие к правому и левому желудочкам. Заканчивается пучок Гиса в толще мышц волокнами Пуркинье.
Синоатриальныи узел является водителем ритма первого порядка. В нем возникают импульсы, которые определяют частоту сокращений сердца. Он генерирует импульсы со средней частотой 70-80 импульсов в 1 мин.
Атриовентрикулярный узел — водитель ритма второго порядка.
Пучок Гиса — водитель ритма третьего порядка.
Волокна Пуркинье — водители ритма четвертого порядка. Частота возбуждения, возникающая в клетках волокон Пуркинье, очень низкая.
В норме атриовентрикулярный узел и пучок Гиса являются только передатчиками возбуждений из ведущего узла к сердечной мышце.
Однако и они обладают автоматизмом, только в меньшей степени, и этот автоматизм проявляется лишь при патологии.
В области синоатриального узла обнаружено значительное число нервных клеток, нервных волокон и их окончаний, которые образуют здесь нервную сеть. К узлам атипической ткани подходят нервные волокна от блуждающих и симпатических нервов.
Возбудимость сердечной мышцы — способность клеток миокарда при действии раздражителя приходить в состояние возбуждения, при котором изменяются их свойства и возникает потенциал действия, а затем сокращение. Сердечная мышца менее возбудима, чем скелетная. Для возникновения возбуждения в ней необходим более сильный раздражитель, чем для скелетной. При этом величина реакции сердечной мышцы не зависит от силы наносимых раздражений (электрических, механических, химических и др.). Сердечная мышца максимально сокращается и на пороговое, и на более сильное по величине раздражение.
Уровень возбудимости сердечной мышцы в разные периоды сокращения миокарда меняется. Так, дополнительное раздражение сердечной мышцы в фазу ее сокращения (систолу) не вызывает нового сокращения даже при действии сверхпорогового раздражителя. В этот период сердечная мышца находится в фазе абсолютной рефрактерности. В конце систолы и начале диастолы возбудимость восстанавливается до исходного уровня — это фаза относительной рефрактерное/пи. За этой фазой следует фаза экзальтации, после которой возбудимость сердечной мышцы окончательно возвращается к исходному уровню. Таким образом, особенностью возбудимости сердечной мышцы является длительный период рефрактерности.
Проводимость сердца — способность сердечной мышцы проводить возбуждение, возникшее в каком-либо участке сердечной мышцы, к другим ее участкам. Возникнув в синоатриальном узле, возбуждение распространяется по проводящей системе на сократительный миокард. Распространение этого возбуждения обусловлено низким электрическим сопротивлением нексусов. Кроме того, проводимости способствуют специальные волокна.
Волны возбуждения проводятся по волокнам сердечной мышцы и атипической ткани сердца с неодинаковой скоростью. Возбуждение по волокнам мышц предсердий распространяется со скоростью 0,8-1 м/с, по волокнам мышц желудочков — 0,8-0,9 м/с, по атипической ткани сердца — 2-4 м/с. При прохождении возбуждения через атриовентрикулярный узел возбуждение задерживается на 0,02- 0,04 с — это атриовентрикулярная задержка, обеспечивающая координацию сокращения предсердий и желудочков.
Сократимость сердца — способность мышечных волокон укорачиваться или изменять свое напряжение. Она реагирует на раздражители нарастающей силы по закону «все или ничего». Сердечная мышца сокращается по типу одиночного сокращения, так как длительная фаза рефрактерности препятствует возникновению тетанических сокращений. В одиночном сокращении сердечной мышцы выделяют: латентный период, фазу укорочения ([[|систола]]), фазу расслабления (диастола). Благодаря способности сердечной мышцы сокращаться только по типу одиночного сокращения сердце выполняет функцию насоса.
Первыми сокращаются мышцы предсердий, затем слой мышц желудочков, обеспечивая тем самым движение крови из полостей желудочков в аорту и легочный ствол.
источник
Физиологические свойства сердечной мышцы и их отличия от скелетной мускулатуры, специфика сердечного сокращения.
Сердечная мышца, как и скелетная, обладает возбудимостью, способностью проводить возбуждение (проводимостью) и сократимостью. Физиологическими особенностями сердечной мышцы является удлиненный рефрактерный период и автоматия.
автоматия. Способность сердца ритмически сокращаться без внешних раздражений, под влиянием импульсов, возникающих в нем самом, называется автоматиейсердца.
В сердце различают рабочую мускулатуру, представленную поперечнополосатой мышцей, в которой клетки кардиомиоциты образуют функциональный синцитий, а также атипические мышечные клетки (пейсмекеры), в которых возникает и проводится возбуждение. Клетки этой ткани богаты протоплазмой, поперечная же исчерченность в них выражена менее четко. К узлам атипической ткани подходят нервные волокна от блуждающих и симпатических нервов.
Скопление атипических клеток образуют проводящую систему сердца (рис.7.12):
В проводящей системе сердца различают:
— синоатриальный узел (СА) –синусовый или предсердно-синусный, или узел Кисс-Фляка (водитель ритма первого порядка), располагающегося на задней стенке правого предсердия у места впадения полых вен;
— атриовентрикулярный узел (АВ) –предсердно-желудочковый или узел Ашоф-Тавара, (водитель ритма второго порядка), находящийся в правом предсердии вблизи перегородки между предсердиями и желудочками;
— пучок Гиса, (водитель ритма третьего порядка), отходит от атриовентрикулярного узла одним стволом, а затем делится на две ножки (ножки пучка Гиса), идущие к правому и левому желудочкам по межжелудочковой перегородке. Заканчивается пучок Гиса в толще мышц волокнами Яна Пуркинье, проводящими возбуждение к мускулатуре желудочков.

Рис. 7.12. Проводящая система сердца (http://medbook.medicina.ru/chapter.php? >
Синоатриальный узел является ведущим в деятельности сердца (главный водитель ритма), в нем генерируются импульсы с частотой 60-80 ударов в минуту, определяющие частоту сокращений сердца. В норме атриовентрикулярный узел и пучок Гиса являются только передатчиками возбуждения из ведущего узла к сердечной мышце. Однако им присуща способность к автоматии, только выражена она в меньшей степени, чем у синоатриального узла, и проявляется лишь в условиях патологии. Атриовентрикулярный узел способен генерировать импульсы с частотой 40-50 в минуту, пучок Гиса – 30-40, волокна Пуркинье – 10-20 импульсов в минуту. Такая последовательность называется убывающим градиентом автоматии.
Возбудимость сердца. В сердце в отличие от других возбудимых тканей имеется значительно выраженный и удлиненный рефрактерный период. Он характеризуется резким снижением возбудимости ткани во время ее активности. Выделяют абсолютный и относительный рефрактерный период (р.п.). Во время абсолютного р.п. какой бы силы не наносили раздражения на сердечную мышцу, она не отвечает на него возбуждением и сокращением. Он соответствует по времени систоле и началу диастолы предсердий и желудочков. Во время относительного р.п. возбудимость сердечной мышцы постепенно возвращается к исходному уровню. В этот период мышца может ответить на раздражитель сильнее порогового. Он обнаруживается во время диастолы предсердий и желудочков.
Сокращение миокарда продолжается около 0.3 с, по времени примерно совпадает с рефрактерной фазой. Следовательно, в период сокращения сердце неспособно реагировать на раздражители. Благодаря выраженному р.п., который длится больше чем период систолы, сердечная мышца неспособна к тетаническому (длительному) сокращению и совершает свою работу по типу одиночного мышечного сокращения.
Сердечная мышца менее возбудима, чем скелетная. Для возникновения возбуждения в сердечной мышце необходимо применить более сильный раздражитель, чем для скелетной. Установлено, что величина реакции сердечной мышцы не зависит от силы наносимых раздражений (электрических, механических, химических и т. д.). Сердечная мышца максимально сокращается и на пороговое, и на более сильное по величине раздражение.
Проводимость сердца. Она обеспечивает распространение возбуждения от клеток водителей ритма по всему миокарду. Распространение возбуждения по сердцу осуществляется электрическим путем.Волны возбуждения проводятся по волокнам сердечной мышцы и атипической ткани сердца с неодинаковой скоростью. Возбуждение по волокнам мышц предсердий распространяется со скоростью 0,8—1,0 м/с, по волокнам мышц желудочков— 0,8—0,9 м/с, по клеткам проводящей системы сердца—0,2 — 4,2 м/с. Наибольшей проводимостью обладают волокна Пуркинье – 3,0 — 4,2 м/с. В атриовентрикулярном узле скорость проведения возбуждения равна 0,2 м/с, в пучке Гиса – 1,0 – 1,5 м/с. Скорость же проведения возбуждения от мышечных волокон предсердий к атриовентрикулярному узлу невысока. Происходящая здесь задержка распространения возбудительного процесса обеспечивает последовательность в работе предсердий и желудочков. Время полного охвата возбуждением желудочков – 0,01 – 0,015 с.
Сократимость сердечной мышцы. Сократимость сердечной мышцы имеет свои особенности. Первыми сокращаются мышцы предсердий, затем — папиллярные мышцы и субэндокардиальный слой мышц желудочков. В дальнейшем сокращение охватывает и внутренний слой желудочков, обеспечивая тем самым движение крови из полостей желудочков в аорту и легочный ствол.
В отличие от скелетных мышц, сократительная реакция которых нарастает при увеличении силы раздражителя сверх пороговой величины, для сердечной мышцы пороговое раздражение является одновременно и максимальным (закон «все или ничего»). Этот обусловлено тем, что волокна миокарда соединены протоплазматическими мостиками.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Да какие ж вы математики, если запаролиться нормально не можете. 8486 —  | 7362 —
| 7362 —  или читать все.
или читать все.
85.95.179.227 © studopedia.ru Не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования. Есть нарушение авторского права? Напишите нам | Обратная связь.
Отключите adBlock!
и обновите страницу (F5)
очень нужно
источник
СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ
Основными свойствами сердечной мышцы, определяющими непрерывное ритмическое сокращение сердца в течение всей жизни организма, являются автоматия, возбудимость, проводимость и сократимость.
Автоматия.Под автоматией понимают способность сердечной мышцы ритмически возбуждаться и сокращаться без каких-иибо внешних по отношению к сердцу воздействий, т.е. без участия нервной системы и гуморальных факторов, доставляемых к сердцу кровью.
Доказательством автоматии сердца послужили следующие наблюдения и эксперименты.
Изолированное сердце, т. е. выведенное из организма и помещенное в питательный раствор, продолжает самопроизвольно сокращаться. Даже разрезанное на кусочки, оно сокращается в том же ритме, что и у здорового животного. Если у животного денер-вировать сердце, т. е. перерезать все нервные стволы, подходящие к сердцу, оно продолжает сокращаться.
На способности работать без воздействия внешних раздражителей основана пересадка сердца. Оживление остановившегося сердца достигается восстановлением спонтанной активности сердца, его автоматии.
В чем причина такого уникального свойства сердца? У большинства беспозвоночных животных автоматия связана с нервными ганглиями, расположенными вблизи сердца, т. е. имеет ней-рогенную природу. У всех же позвоночных животных и у части беспозвоночных автоматия сердца обусловлена не нервными, а мышечными клетками, которые самопроизвольно деполяризуются после каждого потенциала действия. Эти клетки называются пейсмекерами, или «задающими сердечный ритм», или водителями сердечного ритма. Такая теория автоматии сердца называется миогенной.
Способностью к автоматии обладают атипичные мышечные клетки, составляющие проводящую систему сердца.
Ведущую роль в автоматии играет синусный узел. Он обладает наиболее высокой активностью по сравнению с другими участками проводящей системы, частота импульсации в нем наиболее высокая, и он задает определенную частоту сокращения сердца в состоянии физиологического покоя. Такой ритм обычно называют синусным ритмом, а синусный узел — водителем ритма сердца первого порядка.
Если отделить лигатурой синусный узел от предсердий (опыт Станниуса), то обычно сердце останавливается. Однако через некоторое время оно снова начинает сокращаться, но в более редком ритме. Этот ритм «задает» следующий узел проводящей системы — атриовентрикулярный. Более редкие сокращения сердца обусловлены тем, что возбудимость атриовентрикулярного узла меньше, чем синусного. Этот узел называют водителем ритма сердца второго порядка. Если же и атриовентрикулярный узел перестает генерировать возбуждение, то водителем ритма сердца становится пучок Гиса, но его возбудимость еще меньше; пучок Гиса называют водителем ритма третьего порядка.
В обычных условиях атриовентрикулярный узел и пучок Гиса только проводят возбуждение от синусного узла. Их собственная автоматия как бы подавлена главным пейсмекером, и только при развитии патологического процесса, прекращающего функцию
синусного узла, свой ритм навязывают нижележащие узлы. Они являются латентными, или скрытыми, или потенциальными пейсмекерами.
Какова природа автоматии? Методами электрофизиологии установлено, что потенциал действия (ПД) клеток проводящей системы отличается от других мышечных и нервных клеток. Во время расслабления сердца — диастолы — начинается медленно нарастающая деполяризация мембраны, которая затем переходит в фазу быстрой деполяризации (рис. 6.3, А). Фаза реполяризации в пейс-мекерах довольно продолжительная, в пейсмекерах синусного узла она имеет выраженное плато вместо пика потенциала. Сразу после возвращения мембранного потенциала к уровню потенциала покоя снова начинается медленная диастолическая деполяризация мембраны, и когда разность потенциалов между наружной и внутренней поверхностями мембраны уменьшается до определенного критического, или порогового уровня, внезапно возникает новый крутой сдвиг электрического заряда клетки, что свидетельствует о ее возбуждении.
| Ведущий пейсмекер |
 |
| Пороговый потенциал |
| М*едленная\ диастолическая депог ляризация |
| ___ . Макси-___ мольный диастолический потенциал |
 |
| Рабочий миокард |
| Пороговый потенциал L |
| Рис. 6.3. Электрический потенциал пейсмекера (А) и рабочего миокарда (Б) |
Интервал между двумя ПД зависит от длительности медленной диастолической деполяризации, ее величины и порогового уровня сердечного ПД. Если скорость деполяризации уменыиает-
 ся (например, при охлаждении синусного узла), то пороговый уровень деполяризации наступает позднее, частота ПД и сокращений сердца уменьшаются. При возрастании скорости деполяризации мембраны, напротив, пороговый уровень деполяризации возникает раньше и это приводит к учащению возбуждения сердца. Отчасти этим объясняется учащение сердечной деятельности при повышении температуры тела.
ся (например, при охлаждении синусного узла), то пороговый уровень деполяризации наступает позднее, частота ПД и сокращений сердца уменьшаются. При возрастании скорости деполяризации мембраны, напротив, пороговый уровень деполяризации возникает раньше и это приводит к учащению возбуждения сердца. Отчасти этим объясняется учащение сердечной деятельности при повышении температуры тела.
Медленная диастолическая деполяризация обусловлена особенностями ионной проницаемости мембраны пейсмекеров. Как и в других клетках, электрические процессы в мембранах миокарда являются следствием пассивного и активного перемещения ионов натрия и калия через тончайшие каналы (поры) в мембране, проницаемость которых регулируется заряженными частицами — ионами Са 2+ или Мп 2 . Медленная диастолическая деполяризация объясняется тем, что во время реполяризации часть натриевых каналов не инактивируется и осуществляется медленный вход сначала натрия, а затем кальция в мембрану. Когда количество ионов натрия, проникших в клетку, снизит мембранный потенциал до критического уровня, наступает быстрая фаза деполяризации и ПД достигает своего максимального уровня.
В теории об автоматии пейсмекеров еще много неясного, и раскрытие тончайших механизмов электрических процессов, происходящих в сердце, — актуальная задача современной кардиологии.
Возбудимость.Возбудимость — свойство сердечной мышцы переходить в состояние возбуждения под влиянием различных раздражителей.
В естественных условиях раздражителем является ПД, возникающий в синусном узле и распространяющийся по проводящей системе сердца до рабочих кардиомиоцитов. При некоторых заболеваниях сердца раздражение может возникать в других его участках, которые генерируют собственные ПД, и тогда сердечный ритм будет нарушен из-за взаимодействия разных по частоте и фазе ПД. В экспериментах на животных в качестве раздражителей могут быть использованы механические, термические или химические воздействия, если их величина превышает порог возбудимости сердца.
При болезнях сердца, сопровождающихся нарушением сердечного ритма, больным вживляют в сердце миниатюрные электроды, питающиеся от батареек. Импульсы тока подаются непосредственно на сердце и возбуждают в нем ритмические импульсы. При внезапной остановке сердца или нарушении синхронизации отдельных мышечных волокон возможно воздействие на сердце прямо через кожный покров сильным коротким электрическим разрядом напряжением в несколько кВт. Это вызывает одновременное возбуждение всех мышечных волокон, после чего восстанавливается работа сердца.
Во время возбуждения в сердце возникают физико-химические, морфологические и биохимические изменения, которые приводят к сокращению рабочего миокарда. Одними из ранних признаков возбуждения являются активация натриевых каналов и диффузия ионов натрия из межклеточной жидкости через мембрану, что приводит к ее деполяризации и возникновению ПД.
В клетках рабочего миокарда ПД равен 80. 90 мВ, при ПД Ю0. 120мВ медленная диастолическая деполяризация в отличие от пейсмекеров отсутствует. Скорость нарастания деполяризации велика, восходящая часть ПД очень крутая, но реполяризация протекает замедленно, и мембрана остается деполяризованной в течение сотен миллисекунд (см. рис. 6.3, Б).
Таким образом, длительность ПД в миокардиоцитах во много раз больше, чем в других мышечных волокнах. Благодаря этому все мышечные волокна предсердий или желудочков успевают сократиться до того, как какое-либо из этих волокон начнет расслабляться. Поэтому фаза реполяризации продолжается в течение всей систолы. Во время развития ПД возбудимость сердца, как и других возбудимых тканей, изменяется. Во время деполяризации возбудимость сердца резко снижается. Это — фаза абсолютной рефрактерности. Причиной ее является инактивация натриевых каналов, что прекращает поступление новых ионов натрия в мембрану. Если в скелетной мышце абсолютная рефрактерность очень кратковременная, измеряется десятыми долями миллисекунды и заканчивается в начале сокращения мышцы, то в сердце абсолютная невозбудимость продолжается весь период систолы. Практически это означает, что если во время систолы на сердце действует какой-либо раздражитель, даже сверхпороговый, то сердце на него не реагирует. Поэтому в отличие от скелетных мышц сердце не способно к тетаническим сокращениям и защищено от слишком быстрого повторного возбуждения и сокращения. Все сокращения сердечной мышцы одиночные. При очень большой частоте импульсов возбуждения сердце сокращается не на каждый ПД, а на только те из них, которые поступают по окончании абсолютной рефрактерности.
Во время нисходящей фазы реполяризации, которая совпадает с началом расслабления сердечной мышцы, возбудимость сердца начинает восстанавливаться. Это — фаза относительной рефрактерности. Если в начале диастолы на сердце действует какой-либо дополнительный раздражитель, то сердце готово ответить на него новой волной возбуждения. Внеочередное возбуждение и сокращение сердца под действием раздражителя в период относительной рефрактерности называется экстрасистолой.
Если очаг внеочередного возбуждения находится в синусном узле, то это приводит к преждевременному возникновению сер-
Рис. 6.4. Экстрасистола:
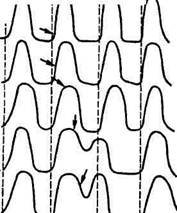 J, 2, 3 — искусственное раздражение наносят во время сокращения желудочков, поэтому эффекта нет (абсолютная рефрактерная фаза); 4, 5, б—раздражение наносят во время расслабления мышцы и вызывают появление экстрасистол, амплитуда которых тем больше, чем позже в период расслабления нанесено раздражение. Это связано с изменением возбудимости мышцы, наступающим после возбуждения; прямые стрелки и пунктирные линии указывают момент появления импульсов в синусном узле, боковые — момент искусственного раздражения сердца
J, 2, 3 — искусственное раздражение наносят во время сокращения желудочков, поэтому эффекта нет (абсолютная рефрактерная фаза); 4, 5, б—раздражение наносят во время расслабления мышцы и вызывают появление экстрасистол, амплитуда которых тем больше, чем позже в период расслабления нанесено раздражение. Это связано с изменением возбудимости мышцы, наступающим после возбуждения; прямые стрелки и пунктирные линии указывают момент появления импульсов в синусном узле, боковые — момент искусственного раздражения сердца
дечного цикла, при этом последовательность сокращений предсердий и желудочков не изменяется. Если же возбуждение возникает в желудочках, то после внеочередного сокращения (экстрасистолы) появляется удлиненная пауза. Интервал между экстрасистолой и следующей (очередной) систолой желудочков называется компенсаторной паузой (рис. 6.4.).
Компенсаторная пауза объясняется тем, что экстрасистола, как и всякое сокращение сердечной мышцы, сопровождается рефрактерной паузой. Очередной импульс, возникающий в синусном узле, приходит в желудочки во время абсолютной рефрактерное™ и не вызывает их сокращения. Новое сокращение наступит лишь в ответ на следующий импульс, когда возбудимость миокарда восстановится.
После относительной рефрактерности в сердце наступает очень короткий период повышенной возбудимости — экзальтации, когда сердце готово ответить даже на подпороговое раздражение.
Проводимость.Проводимость — свойство сердечной мышцы проводить возбуждение.
Как уже сказано, импульс возбуждения (ПД), возникая в пейс-мекерах синусного узла, распространяется сначала на предсердия. В предсердиях, где очень небольшое количество проводящих атипичных мышечных волокон, возбуждение распространяется не только по ним, но и по рабочим кардиомиоцитам. Это объясняет небольшую скорость распространения возбуждения в предсердиях.
Поскольку синусный узел расположен в правом предсердии, а скорость передачи ПД невелика, то возбуждение правого предсер-
дия начинается немного раньше, чем левого. Сокращение же левого и правого предсердий происходит одновременно.
После того как возбуждение охватит мышцы предсердий, они сокращаются, а возбуждение концентрируется и задерживается в атриовентрикулярном узле. Атриовентрикулярная задержка длится до окончания сокращения предсердий, и только после этого возбуждение переходит на пучок Гиса. Таким образом, биологическое значение атриовентрикулярной задержки заключается в обеспечении последовательности сокращений предсердий и желудочков. Одновременное их сокращение иногда бывает при очень серьезной патологии, когда возбуждение возникает не в синусном узле, а в атриовентрикулярном и распространяется в обе стороны от атриовен-трикулярного узла — и в предсердия, и в желудочки. В таком случае наступает резкое нарушение гемодинамики в сердце.
Механизмы атриовентрикулярной задержки не выяснены. Возможно, влияет низкая амплитуда ПД в клетках-пейсмекерах данного узла, сильная натриевая инактивация, большое сопротивление межклеточных контактов.
Далее возбуждение распространяется по пучку Гиса, ножкам пучка Гиса и волокнам Пуркинье. Волокна Пуркинье контактируют с сократительными волокнами миокарда, и возбуждение передается с проводящей системы на рабочие мышцы.
Скорость распространения возбуждения в сердце следующая: от синусного узла до атриовентрикулярного узла — 0,5. 0,8 м/с; в атриовентрикулярном узле — 0,02. 0,05; по проводящей системе желудочков — до 4,0; в сократительной мышце желудочков — 0,4 м/с.
Непосредственная связь проводящей системы сердца с рабочими кардиомиоцитами осуществляется с помощью многочисленных разветвлений волокон Пуркинье. Передача сигналов происходит электрическим путем с небольшой задержкой. Эта задержка возбуждения способствует суммированию импульсов, неодновременно поступающих по волокнам Пуркинье, и обеспечивает лучшую синхронизацию процесса возбуждения рабочего миокарда.
В рабочем миокарде имеются контакты как между торцами, так и боковыми поверхностями волокон. Поэтому возбуждение от основных стволов проводящей системы (ножек пучка Гиса) практически одновременно распространяется на правый и левый желудочки, обеспечивая их одновременное сокращение.
Направление возбуждения внутри желудочков различно у животных разного вида. Так, у собак возбуждение вначале возникает на расстоянии нескольких миллиметров от внутренней поверхности мышечной стенки, а затем переходит к эндокарду и эпикарду. У копытных (у коз) направление распространения возбуждения в толще мышечной стенки меняется много раз, и множество волокон в районах эндокарда, эпикарда и в глубине стенки активируется практически одновременно.
В межжелудочковой перегородке возбуждение начинается в
центральной части и движется к верхушке и атриовентрикулярной
перегородке, причем верхняя часть желудочков активируется поз- ]
же; однако на правой и левой сторонах межжелудочковой перего
родки возбуждение возникает одновременно. j
Особенности распространения возбуждения в сердце имеют значение при анализе электрокардиограммы — записи биотоков сердца.
Сократимость. Сокращение — специфический признак возбуждения сердечной мышцы. Как и в других мышцах, сокращение сердечных мышечных волокон начинается после распространения потенциала действия по поверхности клеточных мембран и является функцией миофибрилл. Сократительная система миофиб-рилл представлена четырьмя белками — актином, миозином, тро-понином и тропомиозином. Сокращение миофибрилл сердца в принципе не отличается от сокращений скелетных мышц согласно теории скольжения протофибрилл Хаксли.
Суть теории Хаксли заключается в скольжении тонких актино-вых нитей в промежутки между толстыми миозиновыми нитями, ; что приводит к укорочению саркомера. При расслаблении мышцы актиновые нити отодвигаются назад, занимая исходное положение. В механизме скольжения актиновых нитей имеет значение кальций, депонированный в саркоплазматическом ретикулуме.
Последовательность электрических и механических процессов при сокращении сердечных мышечных волокон в настоящее время представляется следующим образом. Потенциал действия, возникший на поверхности мембраны мышечного волокна, по поперечным Т-трубочкам, которые являются впячиваниями наружной мембраны, достигает системы поперечных трубочек, соединенных с цистернами саркоплазматического ретикулума. Полости сарко-плазматического ретикулума не сообщаются ни с Т-трубочками, ни с интерстициальной жидкостью и заполнены раствором с высоким содержанием ионов кальция. Полости Т-трубочек имеют такой же состав, что и межклеточная жидкость.
Во время возбуждения активируются натриевые каналы в мембранах Т-трубочек и в миоплазму входят ионы натрия и кальция из межклеточной жидкости. Большая часть входящего кальция не участвует в сокращении миофибрилл, а пополняет его запасы в саркоплазматическом ретикулуме. Под воздействием потенциала действия повышается проницаемость мембраны саркоплазматического ретикулума и ионы кальция вьщеляются из него в миоплазму. Ионы кальция связываются с тропонином, что вызывает конформационные изменения в его молекуле. Сдвиг тропонин-тропомиозинового стержня I обеспечивает взаимодействие нитей актина и миозина (напомним, Щ что в расслабленной мышце актиновые волокна прикрыты молеку- 1 лами тропонина и тропомиозина, образующими комплекс, препятствующий скольжению протофибрилл).
После освобождения актиновых нитей от блокировки тропо-миозиновым комплексом миозиновые головки присоединяются к соответствующему центру актиновых нитей под углом 90°. Затем наступает спонтанный поворот головки на 45°, развивается напряжение и происходит продвижение актиновой нити на один шаг. Эти процессы осуществляются за счет энергии АТФ, причем распад АТФ катализируется актомиозиновым комплексом, обладающим АТФ-азной активностью.
Когда возбуждение прекращается, содержание ионов кальция в миоплазме снижается вследствие работы кальциевого насоса и закачивания кальция в саркоплазматический ретикулум, причем на работу кальциевого насоса также затрачивается энергия АТФ. В результате снижения содержания кальция в миоплазме тропо-миозиновый комплекс защищает активные центры актомиозино-вых нитей. Нити миозина и актина восстанавливают исходное положение, и мышца расслабляется.
Изложенная теория сокращения сердечной мышцы во многом объясняет экспериментальные и клинические наблюдения о влиянии кальция и магния — его антагониста на работу сердца. Известно, что при перфузии изолированного сердца раствором, не содержащим кальция, оно останавливается, а при добавлении кальция в перфузионный раствор сокращения восстанавливаются. Известно также, что сердечные глюкозиды (например, препараты наперстянки) увеличивают проницаемость мембран для кальция и тем самым восстанавливают транспорт кальция между саркоплазматическим ретикулумом, наружной мембраной и миоплазмой.
Согласуется с теорией мышечного сокращения и благоприятное влияние на сердце макроэргических веществ, энергия которых используется не только для механического сокращения, но и для работы ионных насосов — кальциевого и калиево-натриевого.
Сократительные свойства сердечной мышцы несколько отличаются от скелетных. Если скелетная мышца реагирует на раздражение в соответствии с его силой, то сердечная мышца подчиняется закону Боудича «все или ничего». Его суть заключается в том, что на подпороговые раздражения сердце не сокращается («ничего»), а на пороговое раздражение отвечает максимальным сокращением («все»), и увеличение силы раздражителя не приводит к увеличению силы сокращения.
В скелетных мышцах закону «все или ничего» подчиняются отдельные мышечные волокна. Дело в том, что потенциал действия вызывает освобождение кальция из саркоплазматического ретикулума равномерно по всей длине волокна, поэтому оно сокращается полностью. Но в скелетной мышце имеются волокна с разной степенью возбудимости, поэтому при слабом раздражении сокращаются не все волокна и суммарное сокращение оказывается небольшим. В сердечной же мышце волокна рабочего, т. е. сократительного, миокарда соединены межклеточными контактами
(выростами плазматических мембран), что способствует практически одновременному распространению потенциала действия по всей мышце, и она возбуждается и сокращается как единый орган, 1 являясь функциональным синцитием.
Закон Боудича является скорее правилом с определенными ограничениями. При подпороговом раздражении сокращение, действительно, не возникает, но в это время начинается активация натриевых каналов и повышается возбудимость миокардиоцитов. Возникающие местные потенциалы могут суммироваться и вызвать распространяющийся потенциал действия. С другой стороны, сила сокращения сердца, как хорошо известно, непостоянна и может изменяться в различных условиях жизни.
Другая характерная особенность сердечной мышцы заключается в том, что сила сокращения сердца зависит от степени растяжения мышечных волокон во время диастолы, когда полости заполняются кровью. Это — закон Франка — Старлинга. Указанная закономерность объясняется тем, что при растяжении сердца кровью во время диастолы актиновые нити несколько вытягиваются из промежутков между миозиновыми, и при последующем сокращении возрастает число генерирующих силу поперечных мостиков. Кроме того, при растягивании сердечной мышцы в ней повышается сопротивление упругих элементов, и во время сокращения они играют роль «пружины», увеличивая силу сокращения.
Особенно важное значение закон Франка — Старлинга имеет во время усиленной работы сердца, когда возрастает объем крови, поступающей в него во время диастолы. Увеличение силы сокращения приводит к тому, что вся кровь выбрасывается при систоле желудочков в артериальные сосуды, иначе после каждого сокращения в сердце оставалась бы значительная порция крови. При отсутствии большой нагрузки и небольшом объеме кровотока сила сокращения сердца умеренная. Таким образом сердце способно регулировать в известных пределах силу сокращения в зависимости от объема кровотока.
Дата добавления: 2014-11-29 ; Просмотров: 2799 ; Нарушение авторских прав? ;
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
источник